Review Tại sao plasmid là yếu tố giới tính của vi khuẩn
Kinh Nghiệm Hướng dẫn Tại sao plasmid là yếu tố giới tính của vi khuẩn Mới Nhất
Hà Huy Tùng Nguyên đang tìm kiếm từ khóa Tại sao plasmid là yếu tố giới tính của vi khuẩn được Cập Nhật vào lúc : 2022-07-14 18:26:02 . Với phương châm chia sẻ Thủ Thuật Hướng dẫn trong nội dung bài viết một cách Chi Tiết 2022. Nếu sau khi tham khảo tài liệu vẫn ko hiểu thì hoàn toàn có thể lại Comment ở cuối bài để Admin lý giải và hướng dẫn lại nha.
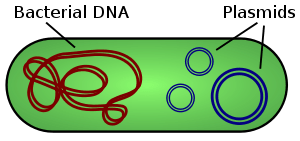
Minh họa về một loại vi khuẩn đã cho tất cả chúng ta biết DNA nhiễm sắc thể và plasmid. Không theo tỷ lệ.
Nội dung chính- Lịch sử [ chỉnh sửa ] Thuộc tính và đặc điểm [ chỉnh sửa ] Phân loại và chủng loại [ chỉnh sửa ] Các vectơ [ chỉnh sửa ] Nhân bản [ chỉnh sửa ] Sản xuất protein [ chỉnh sửa ] Liệu pháp gen [ chỉnh sửa ] Các quy mô bệnh tật [ chỉnh sửa ] Episomes [ chỉnh sửa ] Bảo trì plasmid [ chỉnh sửa ] Các plasmid nấm men [ chỉnh sửa ] Sự phù hợp [ chỉnh sửa ] Phần mềm cho tin sinh học và thiết kế [ chỉnh sửa ] Các bộ sưu tập plasmid [ chỉnh sửa ] Xem thêm [ chỉnh sửa ] Tài liệu tham khảo [ chỉnh sửa ] Further reading[edit]Episomes[edit]External links[edit]
Một plasmid là một phân tử DNA nhỏ trong một tế bào được tách ra khỏi DNA nhiễm sắc thể và hoàn toàn có thể sao chép độc lập. Chúng thường được tìm thấy dưới dạng những phân tử DNA sợi đôi, hình tròn trụ nhỏ ở vi khuẩn; tuy nhiên, plasmid đôi khi xuất hiện trong vi khuẩn cổ và sinh vật nhân chuẩn. Trong tự nhiên, plasmid thường mang những gen hoàn toàn có thể có lợi cho việc sống sót của sinh vật, ví dụ như kháng kháng sinh. Mặc dù nhiễm sắc thể rất lớn và chứa tất cả những thông tin di truyền thiết yếu để sống trong điều kiện thông thường, nhưng plasmid thường rất nhỏ và chỉ chứa những gen tương hỗ update hoàn toàn có thể hữu ích cho sinh vật trong một số trong những tình huống hoặc điều kiện rõ ràng. Các plasmid tự tạo được sử dụng rộng rãi như những vectơ trong nhân bản phân tử, phục vụ cho việc sao chép những chuỗi DNA tái tổ hợp trong khung hình vật chủ. Trong phòng thí nghiệm, plasmid hoàn toàn có thể được đưa vào tế bào thông qua biến hóa.
Plasmid được xem là bản sao đơn vị DNA hoàn toàn có thể sao chép tự chủ trong một vật chủ thích hợp. Tuy nhiên, những plasmid, in như virut, thường không được phân loại là sự việc sống. [1] Các plasmid được truyền từ vi khuẩn này sang vi khuẩn khác (thậm chí của một loài khác) đa phần thông qua phối hợp. [2] Sự quy đổi vật chủ này sang vật chủ di truyền là một cơ chế chuyển gen ngang, và plasmid được xem là một phần của vận động. Không in như virut (bao bọc vật liệu di truyền của chúng trong lớp vỏ protein bảo vệ gọi là capsid), plasmid là DNA "trần trụi" và không mã hóa những gen thiết yếu để bọc vật liệu di truyền để chuyển sang vật chủ mới. Tuy nhiên, một số trong những loại plasmid mã hóa pilus "sex" phối hợp thiết yếu cho việc chuyển giao của chúng. Kích thước của plasmid thay đổi từ 1 đến hơn 200 kbp, [3] và số lượng plasmid giống hệt nhau trong một tế bào hoàn toàn có thể xấp xỉ từ một đến Hàng trăm trong một số trong những trường hợp.
Mối quan hệ giữa vi khuẩn và DNA plasmid không ký sinh cũng không tương hỗ, chính bới mỗi loại ý niệm sự hiện hữu của một loài độc lập sống trong trạng thái bất lợi hoặc giao tế với sinh vật chủ. Thay vào đó, plasmid đáp ứng một cơ chế chuyển gen ngang trong quần thể vi khuẩn và thường đáp ứng một lợi thế tinh lọc trong một trạng thái môi trường tự nhiên thiên nhiên nhất định. Plasmid hoàn toàn có thể mang gen kháng kháng sinh tự nhiên trong môi trường tự nhiên thiên nhiên đối đầu đối đầu hoặc protein được sản xuất hoàn toàn có thể hoạt động và sinh hoạt giải trí như độc tố trong trường hợp tương tự hoặc được cho phép sinh vật sử dụng những hợp chất hữu cơ đặc biệt có lợi khi dinh dưỡng khan hiếm. [4]
Lịch sử [ chỉnh sửa ]
Thuật ngữ plasmid được ra mắt vào năm 1952 bởi nhà sinh học phân tử người Mỹ Joshua Lederberg để đề cập đến "bất kỳ yếu tố quyết định di truyền ngoại bào nào". thuật ngữ được sử dụng sớm gồm có bất kỳ vật liệu di truyền vi khuẩn nào tồn tại ngoại bào trong ít nhất một phần của chu kỳ luân hồi sao chép của nó, nhưng vì mô tả đó gồm có virus vi khuẩn, khái niệm plasmid đã được tinh chế theo thời gian để gồm có những yếu tố di truyền sinh sản tự chủ. [6] , người ta đã quyết định rằng thuật ngữ plasmid nên được sử dụng làm thuật ngữ cho yếu tố di truyền ngoại bào, [7] và để disti Đánh lừa nó khỏi virus, định nghĩa được thu hẹp thành những yếu tố di truyền tồn tại độc quyền hoặc đa phần bên phía ngoài nhiễm sắc thể và hoàn toàn có thể sao chép một cách tự chủ. [6]
Thuộc tính và đặc điểm [ chỉnh sửa ]
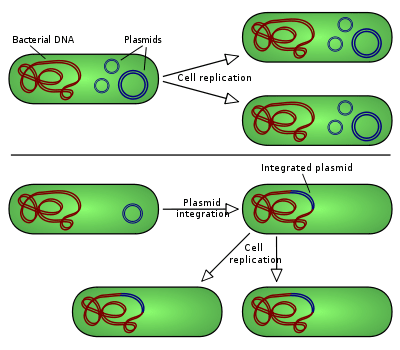
Các loại tích hợp plasmid vào vi khuẩn chủ: Các plasmid không tích hợp sao chép như với trường hợp số 1, trong khi những episome, ví dụ thấp hơn, hoàn toàn có thể tích hợp vào nhiễm sắc thể của vật chủ.
Để những plasmid sao chép độc lập trong một tế bào, chúng phải sở hữu một đoạn DNA hoàn toàn có thể đóng vai trò là nguồn gốc của sự việc sao chép. Đơn vị tự sao chép, trong trường hợp này là plasmid, được gọi là bản sao. Một bản sao vi khuẩn điển hình hoàn toàn có thể gồm có một số trong những yếu tố, ví dụ như gen cho protein khởi tạo sao chép đặc hiệu plasmid (Rep), những đơn vị lặp lại được gọi là iterons, hộp DnaA và một khu vực giàu AT liền kề. [6] Các plasmid nhỏ hơn sử dụng của những enzyme sao chép của vật chủ để tạo ra những bản sao của chúng, trong khi những plasmid to hơn hoàn toàn có thể mang những gen đặc trưng cho việc sao chép của những plasmid đó. Một vài loại plasmid cũng hoàn toàn có thể chèn vào nhiễm sắc thể chủ và những plasmid tích hợp này đôi khi được gọi là episome trong prokaryote. [8]
Plasmid hầu như luôn mang ít nhất một gen. Nhiều gen được mang bởi một plasmid có lợi cho những tế bào chủ, ví dụ: được cho phép tế bào chủ tồn tại trong một môi trường tự nhiên thiên nhiên hoàn toàn có thể gây chết người hoặc hạn chế sự phát triển. Một số gen này mã hóa những tính trạng kháng kháng sinh hoặc kháng với sắt kẽm kim loại nặng, trong khi những gen khác hoàn toàn có thể tạo ra những yếu tố độc lực được cho phép vi khuẩn xâm chiếm vật chủ và vượt qua sự phòng vệ của nó, hoặc có hiệu suất cao trao đổi chất rõ ràng được cho phép vi khuẩn sử dụng một chất dinh dưỡng rõ ràng, gồm hoàn toàn có thể làm suy giảm recalcitrant hoặc những hợp chất hữu cơ độc hại. [6] Plasmid cũng hoàn toàn có thể đáp ứng cho vi khuẩn kĩ năng cố định và thắt chặt nitơ. Tuy nhiên, một số trong những plasmid không còn công dụng quan sát được trên kiểu hình của tế bào chủ hoặc quyền lợi của nó đối với tế bào chủ không thể xác định được và những plasmid này được gọi là plasmid mật mã. [9]
rất khác nhau rất nhiều trong tính chất vật lý của tớ. Kích thước của chúng hoàn toàn có thể xấp xỉ từ những plasmid nhỏ rất nhỏ dưới 1 cặp kilobase (Kbp), đến những megapixel rất lớn của một vài cặp megabase (Mbp). Ở đầu cuối, rất ít hoàn toàn có thể phân biệt giữa một megapixel và một minichromosome. Plasmid thường có hình tròn trụ, nhưng những ví dụ về plasmid tuyến tính cũng khá được nghe biết. Các plasmid tuyến tính này đòi hỏi những cơ chế chuyên biệt để tái tạo những đầu của chúng. [6]
Các plasmid hoàn toàn có thể có trong một tế bào riêng lẻ với số lượng rất khác nhau, từ một đến vài trăm. Số lượng bản sao thông thường của plasmid hoàn toàn có thể được tìm thấy trong một tế bào được gọi là số bản sao Plasmid và được xác định bằng phương pháp khởi đầu sao chép được quy định và kích thước của phân tử. Các plasmid to hơn có xu hướng có số lượng bản sao thấp hơn. [8] Các plasmid số lượng bản sao thấp chỉ tồn tại dưới dạng một hoặc một vài bản sao trong mỗi vi khuẩn, khi phân chia tế bào, có rủi ro tiềm ẩn tiềm ẩn bị mất ở một trong những vi khuẩn phân tách. Các plasmid sao chép đơn như vậy có những khối mạng lưới hệ thống nỗ lực dữ thế chủ động phân phối một bản sao cho tất cả những tế bào con. Các khối mạng lưới hệ thống này, gồm có khối mạng lưới hệ thống parABS và khối mạng lưới hệ thống parMRC, thường được gọi là khối mạng lưới hệ thống phân vùng hoặc hiệu suất cao phân vùng của một plasmid.
Phân loại và chủng loại [ chỉnh sửa ]
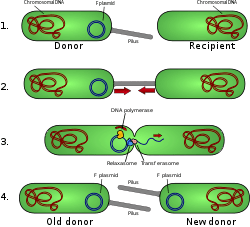
Tổng quan về sự phối hợp của vi khuẩn
Máy vi tính điện tử của một plasmid DNA của vi khuẩn (đoạn nhiễm sắc thể).
Plasmid hoàn toàn có thể được phân loại cách Plasmid hoàn toàn có thể được phân loại thành plasmid phối hợp và plasmid không phối hợp. Các plasmid phối hợp chứa một bộ gen chuyển hoặc tra gen thúc đẩy sự phối hợp tình dục Một trong những tế bào rất khác nhau. [8] Trong quá trình phối hợp phức tạp, plasmid hoàn toàn có thể được chuyển từ vi khuẩn này sang vi khuẩn khác thông qua pili được mã hóa bởi một số trong những vi khuẩn. những gen tra (xem hình). [10] Các plasmid không phối hợp không hoàn toàn có thể khởi đầu phối hợp, do đó chúng chỉ hoàn toàn có thể được chuyển với sự tương hỗ của những plasmid phối hợp. Một lớp plasmid trung gian hoàn toàn có thể vận động được và chỉ mang một tập hợp con những gen thiết yếu để chuyển. Chúng hoàn toàn có thể ký sinh một plasmid phối hợp, chuyển với tần số cao chỉ khi xuất hiện.
Plasmid cũng hoàn toàn có thể được phân loại thành những nhóm không tương thích. Một vi khuẩn hoàn toàn có thể chứa nhiều chủng loại plasmid rất khác nhau, nhưng những plasmid rất khác nhau chỉ hoàn toàn có thể tồn tại trong một tế bào vi khuẩn nếu chúng tương thích. Nếu hai plasmid không tương thích, cái này hoặc cái kia sẽ nhanh gọn bị mất khỏi tế bào. Do đó, những plasmid rất khác nhau hoàn toàn có thể được gán cho những nhóm không tương thích rất khác nhau tùy thuộc vào việc chúng hoàn toàn có thể cùng tồn tại với nhau hay là không. Các plasmid không tương thích (thuộc cùng một nhóm không tương thích) thường chia sẻ cùng một cơ chế sao chép hoặc phân vùng và do đó không thể được giữ lại trong cùng một ô. [11] [12]
] Một cách khác để phân loại plasmid là theo hiệu suất cao. Có năm lớp chính:
- F-plasmids sinh sản, chứa tra gen. Chúng hoàn toàn có thể phối hợp và dẫn đến sự biểu lộ của pili giới tính.
Các plasmid kháng (R), có chứa những gen đáp ứng kĩ năng chống lại kháng sinh hoặc chất độc. Trong lịch sử được gọi là những yếu tố R, trước khi bản chất của plasmid được hiểu.
Plasmid col, chứa những gen mã hóa cho những vi khuẩn, protein hoàn toàn có thể tiêu diệt những vi khuẩn khác.
Các plasmid thoái hóa, được cho phép tiêu hóa những chất không bình thường, ví dụ toluene và axit salicylic.
Plasmid độc lực, biến vi khuẩn thành mầm bệnh.
Plasmid hoàn toàn có thể thuộc về nhiều hơn nữa một trong những nhóm hiệu suất cao này.
Các vectơ [ chỉnh sửa ]
Các plasmid được sản xuất tự tạo hoàn toàn có thể được sử dụng làm vectơ trong kỹ thuật di truyền. Các plasmid này đóng vai trò là công cụ quan trọng trong phòng thí nghiệm di truyền và công nghệ tiên tiến sinh học, nơi chúng thường được sử dụng để nhân bản và khuếch đại (tạo ra nhiều bản sao) hoặc thể hiện những gen đặc biệt. [13] Nhiều loại plasmid có sẵn trên thị trường cho những mục tiêu sử dụng đó. Các gen được sao chép thường được đưa vào một plasmid thường chứa một số trong những tính năng để sử dụng. Chúng gồm có một gen liên quan đến tính kháng với nhiều chủng loại kháng sinh đặc biệt (ampicillin được sử dụng thường xuyên nhất cho những chủng vi khuẩn), nguồn gốc của sự việc sao chép để được cho phép những tế bào vi khuẩn sao chép DNA plasmid và một vị trí thích hợp để nhân bản (gọi là vị trí nhân bản ).
Sự mất ổn định cấu trúc DNA hoàn toàn có thể được định nghĩa là một chuỗi những sự kiện tự phát mà đỉnh điểm là sự việc sắp xếp lại không lường trước, mất hoặc thu được vật liệu di truyền. Các sự kiện như vậy thường được kích hoạt bởi sự hoán vị của những yếu tố di động hoặc bởi sự hiện hữu của những yếu tố tạm bợ như cấu trúc không chính tắc (không B). Các khu vực phụ kiện liên quan đến xương sống của vi khuẩn hoàn toàn có thể tham gia vào một loạt những hiện tượng kỳ lạ mất ổn định cấu trúc. Các chất xúc tác nổi tiếng về sự mất ổn định di truyền gồm có lặp lại trực tiếp, đảo ngược và song song, được biết là hay thấy trong một số trong những lượng lớn những vectơ nhân bản và biểu lộ có sẵn trên thị trường. [14] Trình tự chèn cũng hoàn toàn có thể ảnh hưởng nghiêm trọng đến hiệu suất cao và năng suất plasmid. để xóa và sắp xếp lại, kích hoạt, điều hòa giảm hoặc bất hoạt biểu lộ gen lân cận. [15] Do đó, việc giảm hoặc vô hiệu hoàn toàn những chuỗi xương sống không mã hóa bên phía ngoài sẽ làm giảm đáng kể xu hướng cho những sự kiện đó xảy ra, và do đó, tái tổ hợp tổng thể tiềm năng của plasmid. [16][17]
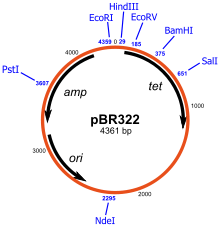
Một đại diện sơ đồ của plasmid pBR322, một trong những plasmid đầu tiên được sử dụng rộng rãi như một vector nhân bản. Trên sơ đồ plasmid là những gen được mã hóa ( amp và tet cho kĩ năng kháng ampicillin và tetracycline tương ứng), nguồn gốc sao chép của nó ( ori ) và nhiều hạn chế rất khác nhau những trang web (được chỉ định bằng màu xanh lam).
Nhân bản [ chỉnh sửa ]
Plasmid là những vectơ nhân bản được sử dụng phổ biến nhất của vi khuẩn. [18] được chèn vào, ví dụ như một trang web nhân bản hoặc polylinker có một số trong những vị trí hạn chế thường được sử dụng mà những đoạn DNA hoàn toàn có thể được nối. Sau khi gen quan tâm được chèn vào, những plasmid được đưa vào vi khuẩn bằng một quá trình gọi là biến hóa. Các plasmid này chứa một chất đánh dấu hoàn toàn có thể lựa chọn, thường là gen kháng kháng sinh, giúp vi khuẩn hoàn toàn có thể sống sót và sinh sôi nảy nở trong môi trường tự nhiên thiên nhiên tăng trưởng tinh lọc có chứa nhiều chủng loại kháng sinh đặc biệt. Các tế bào sau khi biến hóa được tiếp xúc với môi trường tự nhiên thiên nhiên tinh lọc và chỉ có những tế bào chứa plasmid mới hoàn toàn có thể tồn tại. Theo cách này, kháng sinh hoạt động và sinh hoạt giải trí như một bộ lọc để chỉ chọn vi khuẩn chứa DNA plasmid. Vectơ cũng hoàn toàn có thể chứa những gen đánh dấu hoặc gen phóng viên khác để tạo điều kiện lựa chọn plasmid với nhân bản vô tính. Vi khuẩn có chứa plasmid sau đó hoàn toàn có thể được phát triển với số lượng lớn, được thu hoạch và plasmid quan tâm sau đó hoàn toàn có thể được phân lập bằng những phương pháp điều chế plasmid rất khác nhau.
Một vectơ nhân bản plasmid thường được sử dụng để nhân bản những đoạn DNA lên đến mức 15 kbp. [19] Để sao chép DNA dài hơn thế nữa, phage lambda với gen lysogeny đã bị xóa, cosmids, nhiễm sắc thể tự tạo của vi khuẩn hoặc nhiễm sắc thể tự tạo được sử dụng.
Sản xuất protein [ chỉnh sửa ]
Một hiệu suất cao chính khác của plasmid là tạo ra một lượng lớn protein. Trong trường hợp này, những nhà nghiên cứu và phân tích phát triển vi khuẩn có chứa một plasmid chứa gen quan tâm. Giống như vi khuẩn tạo ra protein để tạo ra tính kháng kháng sinh, nó cũng hoàn toàn có thể được tạo ra để tạo ra một lượng lớn protein từ gen được chèn vào. Đây là một cách rẻ tiền và thuận tiện và đơn giản để sản xuất hàng loạt protein, mã gen, ví dụ, insulin.
Liệu pháp gen [ chỉnh sửa ]
Plasmid cũng hoàn toàn có thể được sử dụng để chuyển gen vào tế bào người như thể phương pháp điều trị tiềm năng trong liệu pháp gen để nó hoàn toàn có thể biểu lộ protein thiếu trong tế bào . Một số kế hoạch của liệu pháp gen đòi hỏi phải chèn những gen trị liệu vào những vị trí tiềm năng nhiễm sắc thể được chọn trước trong bộ gen của con người. Các vectơ plasmid là một trong nhiều phương pháp hoàn toàn có thể được sử dụng cho mục tiêu này. Các hạt nhân ngón tay kẽm (ZFN) đáp ứng một phương pháp để phá vỡ chuỗi kép rõ ràng tại chỗ đối với bộ gen DNA và gây ra sự tái hợp tương đồng. Plasmid mã hóa ZFN hoàn toàn có thể giúp đưa một gen trị liệu đến một vị trí rõ ràng để tránh tổn thương tế bào, đột biến gây ung thư hoặc đáp ứng miễn dịch. [20]
Các quy mô bệnh tật [ chỉnh sửa ]
Plasmid trong lịch sử được sử dụng để thiết kế di truyền những tế bào gốc phôi của chuột để tạo ra những quy mô bệnh di truyền chuột. Hiệu quả hạn chế của những kỹ thuật nhờ vào plasmid đã ngăn cản việc sử dụng chúng trong việc tạo ra những quy mô tế bào người đúng chuẩn hơn. Tuy nhiên, sự phát triển trong những kỹ thuật tái tổ hợp virus liên quan đến Adeno và những hạt nhân ngón tay Kẽm đã được cho phép tạo ra một thế hệ mới của những quy mô bệnh tự tạo ở người.
Episomes [ chỉnh sửa ]
Thuật ngữ episome được ra mắt bởi François Jacob và Élie Wollman vào năm 1958 để đề cập đến vật liệu di truyền ngoài nhiễm sắc thể hoàn toàn có thể sao chép một cách tự động hoặc được tích hợp vào nhiễm sắc thể. [21][22] Kể từ khi thuật ngữ này được ra mắt, tuy nhiên, việc sử dụng nó đã thay đổi, vì plasmid đã trở thành thuật ngữ ưa thích để sao chép DNA ngoại bào tự động. Tại một hội nghị chuyên đề năm 1968 ở Luân Đôn, một số trong những người dân tham gia nhận định rằng thuật ngữ episome bị bỏ rơi, tuy nhiên những người dân khác tiếp tục sử dụng thuật ngữ này với một sự thay đổi trong ý nghĩa. [23]
Ngày nay, một số trong những tác giả sử dụng episome trong toàn cảnh prokaryote để chỉ một plasmid hoàn toàn có thể tích hợp vào nhiễm sắc thể. Các plasmid tích hợp hoàn toàn có thể được sao chép và duy trì ổn định trong một tế bào qua nhiều thế hệ, nhưng luôn ở một quá trình nào đó chúng tồn tại như một phân tử plasmid độc lập. [25] Trong toàn cảnh của sinh vật nhân chuẩn, thuật ngữ episomes được sử dụng để nghĩa là một phân tử DNA tuần hoàn khép kín không tích hợp hoàn toàn có thể được sao chép trong nhân. [26][27] Virus là những ví dụ phổ biến nhất về điều này, ví dụ như herpesvirus, adenovirus và polyomavirus, nhưng một số trong những là plasmid. Các ví dụ khác gồm có những đoạn nhiễm sắc thể không bình thường, ví dụ như nhiễm sắc thể phút, hoàn toàn có thể phát sinh trong quá trình khuếch đại gen tự tạo hoặc trong những quá trình bệnh lý (ví dụ: biến hóa tế bào ung thư). Các episome ở sinh vật nhân chuẩn hoạt động và sinh hoạt giải trí tương tự như plasmid ở sinh vật nhân sơ ở chỗ DNA được duy trì ổn định và được sao chép với tế bào chủ. Các biểu lộ virus tế bào chất (như trong nhiễm trùng poxvirus) cũng hoàn toàn có thể xảy ra. Một số episome, ví dụ như herpesvirus, sao chép theo cơ chế vòng tròn lăn, tương tự như virus phage của vi khuẩn. Những người khác sao chép thông qua cơ chế sao chép hai chiều ( Loại theta plasmid). Trong cả hai trường hợp, episome vẫn tách biệt về mặt vật lý với nhiễm sắc thể tế bào chủ. Một số virus ung thư, gồm có virus Epstein-Barr và herpesvirus liên quan đến sarcoma của Kaposi, được duy trì dưới dạng những episome tiềm ẩn, nhiễm sắc thể trong những tế bào ung thư, trong đó những virus biểu lộ gen gây ung thư thúc đẩy sự tăng sinh tế bào ung thư. Trong ung thư, những episome này sao chép thụ động cùng với nhiễm sắc thể chủ khi tế bào phân chia. Khi những episome virus này khởi đầu sao chép lylic để tạo ra nhiều hạt virus, chúng thường kích hoạt những cơ chế bảo vệ miễn dịch bẩm sinh tế bào giết chết tế bào chủ.
Bảo trì plasmid [ chỉnh sửa ]
Một số plasmid hoặc sever vi sinh vật gồm có khối mạng lưới hệ thống gây nghiện hoặc khối mạng lưới hệ thống tiêu diệt sau khi tổng hợp (PSK), ví dụ như giết chết / ức chế sever ) khối mạng lưới hệ thống của plasmid R1 trong Escherichia coli . [28] Biến thể này tạo ra cả chất độc tồn tại lâu và thuốc giải độc ngắn. Một số loại khối mạng lưới hệ thống nghiện plasmid (độc tố / antitoxin, khối mạng lưới hệ thống ORT nhờ vào chuyển hóa) đã được mô tả trong tài liệu [29] và được sử dụng trong những ứng dụng công nghệ tiên tiến sinh học (lên men) hoặc y sinh (liệu pháp vắc-xin). Các tế bào con gái giữ lại một bản sao của plasmid tồn tại, trong khi một tế bào con không thừa hưởng plasmid sẽ chết hoặc bị tụt giảm độ tăng trưởng do chất độc còn sót lại từ tế bào cha. Cuối cùng, năng suất tổng thể hoàn toàn có thể được nâng cao.
trái lại, hầu như tất cả những plasmid được sử dụng công nghệ tiên tiến sinh học (như pUC18, pBR322 và vec tơ dẫn xuất) không chứa những khối mạng lưới hệ thống nghiện độc tố-kháng độc tố và do đó nên phải được giữ dưới áp lực kháng sinh để tránh mất plasmid.
Các plasmid nấm men [ chỉnh sửa ]
Nấm men tự nhiên chứa nhiều plasmid rất khác nhau. Đáng để ý quan tâm trong số đó là 2 plasmid – những plasmid tròn nhỏ thường được sử dụng cho kỹ thuật di truyền của nấm men – và những plasmid pGKL tuyến tính từ Kluyveromyces lactis phụ trách cho kiểu hình kẻ giết người. [30] 19659003] Các loại plasmid khác thường liên quan đến những vectơ nhân bản men gồm có:
- Plasmid tổng hợp nấm men (YIp) những vec tơ nấm men nhờ vào sự tích hợp vào nhiễm sắc thể của vật chủ để tồn tại và sao chép, và thường được sử dụng khi nghiên cứu và phân tích hiệu suất cao của gen solo hoặc khi gen độc hại. Cũng được link với gen URA3, mã hóa một enzyme liên quan đến sinh tổng hợp nucleotide pyrimidine (T, C);
Plasmid sao chép men (YRp) vận chuyển một chuỗi DNA nhiễm sắc thể gồm có một chuỗi nhiễm sắc thể nguồn gốc nhân rộng. Các plasmid này kém ổn định hơn, vì chúng hoàn toàn có thể bị mất trong quá trình nảy chồi.
Như đã nói ở trên, plasmid thường được sử dụng để tinh chế một chuỗi rõ ràng, vì chúng hoàn toàn có thể thuận tiện và đơn giản được tinh chế khỏi phần còn sót lại của cục gen. Để sử dụng làm vectơ và để nhân bản phân tử, plasmid thường nên phải được phân lập.
Có một số trong những phương pháp để phân lập DNA plasmid khỏi vi khuẩn, những nguyên mẫu trong số đó là miniprep và maxiprep / Bulkprep . hoàn toàn có thể được sử dụng để nhanh gọn tìm ra liệu plasmid có đúng trong bất kỳ bản sao nào của vi khuẩn hay là không. Sản lượng là một lượng nhỏ DNA plasmid không tinh khiết, đủ để phân tích bằng phương pháp tiêu hóa hạn chế và cho một số trong những kỹ thuật nhân bản.
Sau này, khối lượng huyền phù vi khuẩn to hơn nhiều được phát triển từ đó hoàn toàn có thể thực hiện sẵn sàng sẵn sàng maxi. Về bản chất, đây là một miniprep được nhân rộng theo sau là thanh lọc tương hỗ update. Điều này dẫn đến số lượng tương đối lớn (vài trăm microgam) DNA plasmid rất tinh khiết.
Trong thời gian mới gần đây, nhiều bộ dụng cụ thương mại đã được tạo ra để thực hiện chiết xuất plasmid ở nhiều quy mô, độ tinh khiết và mức độ tự động hóa rất khác nhau. Các Thương Mại hoàn toàn có thể sẵn sàng sẵn sàng DNA plasmid với niêm yết dưới 300 đô la / mg với số lượng milligram và 15 đô la / mg về số lượng gram (đầu năm 2007 [update]).
Sự phù hợp [ chỉnh sửa ]
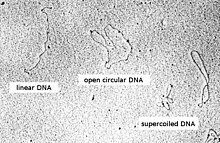
DNA plasmid hoàn toàn có thể xuất hiện ở một trong năm sự phù hợp, (với kích thước nhất định) chạy ở tốc độ rất khác nhau trong gel trong quá trình điện di. Sự phù hợp được liệt kê dưới đây theo thứ tự di động điện di (tốc độ cho một điện áp ứng dụng nhất định) từ chậm nhất đến nhanh nhất có thể:
- DNA có hình tròn trụ mở DNA có một sợi bị cắt.
Thông tư thư giãn DNA hoàn toàn nguyên vẹn với cả hai sợi không biến thành cắt, nhưng đã bị cắt bỏ (siêu vô hiệu). Điều này hoàn toàn có thể được quy mô hóa bằng phương pháp để một sợi dây mở rộng xoắn giãn ra và thư giãn và sau đó cắm nó vào chính nó.
Tuyến tính DNA có kết thúc miễn phí, vì cả hai sợi đã bị cắt hoặc do DNA bị tuyến tính in vivo . Điều này hoàn toàn có thể được quy mô hóa bằng một dây nối điện không được cắm vào chính nó.
Supercoiled (hoặc DNA khép kín hóa trị ) DNA hoàn toàn nguyên vẹn với cả hai sợi không biến thành cắt một xoắn không thể thiếu, dẫn đến một hình thức nhỏ gọn. Điều này hoàn toàn có thể được quy mô hóa bằng phương pháp xoắn một sợi dây mở rộng và sau đó cắm nó vào chính nó.
DNA bị biến tính siêu lọc DNA in như DNA siêu tải nhưng có những vùng không ghép đôi; điều này hoàn toàn có thể xảy ra do độ kiềm quá mức trong quá trình điều chế plasmid.
Tốc độ di tán của những đoạn tuyến tính nhỏ tỷ lệ thuận với điện áp đặt ở điện áp thấp. Ở điện áp cao hơn, những mảnh to hơn di tán với tốc độ tăng liên tục nhưng rất khác nhau. Do đó, độ phân giải của gel giảm khi điện áp tăng.
Ở một điện áp thấp, chỉ định, tốc độ di tán của những đoạn DNA tuyến tính nhỏ là một hàm của độ dài của chúng. Các đoạn tuyến tính lớn (trên 20 kb hoặc hơn) di tán với tốc độ cố định và thắt chặt nhất định bất kể chiều dài. Điều này là vì những phân tử 'hồi sinh', với phần lớn phân tử theo đầu cuối thông qua ma trận gel. Các tiêu hóa hạn chế thường được sử dụng để phân tích những plasmid tinh khiết. Những enzyme này đặc biệt phá vỡ DNA ở một số trong những trình tự ngắn nhất định. Các mảnh tuyến tính thu được tạo thành 'dải' sau khi điện di gel. Có thể làm sạch một số trong những đoạn nhất định bằng phương pháp cắt những dải ra khỏi gel và hòa tan gel để giải phóng những đoạn DNA.
Do cấu trúc ngặt nghèo, DNA siêu tải di tán nhanh hơn qua gel so với DNA tuyến tính hoặc vòng tròn mở.
Phần mềm cho tin sinh học và thiết kế [ chỉnh sửa ]
Việc sử dụng plasmid như một kỹ thuật trong sinh học phân tử được tương hỗ bởi phần mềm tin sinh học. Các chương trình này ghi lại trình tự DNA của những vectơ plasmid, giúp Dự kiến những vị trí cắt của những enzyme cắt số lượng giới hạn và lập kế hoạch thao tác. Ví dụ về những gói phần mềm xử lý những map plasmid là ApE, Clone Manager, GeneCon cảnKit, Geneious, Genome Compiler, LabGenius, Lasergene, MacVector, pDraw32, serial Cloner, VectorFriends, Vector NTI và WebDSV. Những phần mềm này giúp tiến hành toàn bộ những thí nghiệm trong silico trước khi thực hiện những thí nghiệm ướt. [31]
Các bộ sưu tập plasmid [ chỉnh sửa ]
Nhiều plasmid đã được tạo ra trong nhiều năm và những nhà nghiên cứu và phân tích đã đưa ra plasmid cho cơ sở tài liệu plasmid ví dụ như những tổ chức phi lợi nhuận Addgene và BCCM / LMBP. Người ta hoàn toàn có thể tìm và yêu cầu những plasmid từ những cơ sở tài liệu đó để tiếp tục nghiên cứu và phân tích. Nhà nghiên cứu và phân tích cũng thường tải lên những chuỗi plasmid trong cơ sở tài liệu NCBI. Sử dụng cơ sở tài liệu NCBI, những chuỗi plasmid rõ ràng hoàn toàn có thể được tra cứu.
Xem thêm [ chỉnh sửa ]
Tài liệu tham khảo [ chỉnh sửa ]
^ Sinkovics, J; Harvestath J; Horak A. (1998). "Nguồn gốc và sự phát triển của virus (một đánh giá)". Acta Microbiologica et Immunologica Hungarica . 45 (3ùn4): 349 Tắt90. PMID 9873943. ^ Smillie, Chris; Garcillán-Barcia, M. Pilar; Francia, M. Victoria; Rocha, Eduardo P. C.; Cruz, Fernando de la (ngày một tháng 9 năm 2010). "Tính di động của Plasmid". Đánh giá vi sinh và phân tử sinh học . 74 (3): 434 phép52. doi: 10.1128 / MMBR.00020-10. ISSN 1092-2172. PMC 2937521 . PMID 20805406. ^ Thomas, Christopher M; Mùa hè, David (2008). Plasmid vi khuẩn . Bách khoa toàn thư về khoa học sự sống . doi: 10.1002 / YAM470015902.a0000468.pub2. Sê-ri 980-0470016176. ^ Wolfgang Schumann (2008). "Chương 1 – Escherichia coli Nhân bản và biểu lộ vectơ". Trong Georg Lipps. Plasmid: Nghiên cứu hiện tại và xu hướng tương lai . Caister Báo chí học thuật. trang 1 Tiếng2. ISBN 97-1-904455-35-6. CS1 duy trì: Sử dụng tham số sửa đổi và biên tập viên (link) ^ Lederberg J (1952). "Di truyền tế bào và cộng sinh di truyền". Vật lý trị liệu. Rev . 32 (4): 403 Chiếc430. CiteSeerX 10.1.1.458.985 . doi: 10.1152 / Physrev.1952.32.4.403. PMID 13003535. ^ a b c e Finbarr Hayes (2003). "Chương 1 – Chức năng và tổ chức của Plasmid". Trong Nicola Casali, Andrew Presto. E. Coli Plasmid vectơ: Phương pháp và ứng dụng . Các phương pháp trong sinh học phân tử. 235 . Báo chí Humana. tr.115. ISBN 97-1-58829-151-6. CS1 duy trì: Sử dụng tham số trình sửa đổi (link) ^ Stanley Falkow. "Bộ gen của vi sinh vật: Đứng trên vai người khổng lồ". Thương Hội Vi sinh học . ^ a b c A. Nâu (2010). "Chương 2 – Các vectơ cho nhân bản gen: Plasmid và vi khuẩn". Phân tích gen và phân tích DNA: Giới thiệu (tái bản lần thứ 6). Wiley-Blackwell. Sê-ri 980-1405181730. ^ David Summers (1996). "Chương 1 – Chức năng và tổ chức của Plasmid". Sinh học của Plasmid . Wiley-Blackwell; Ấn bản đầu tiên. trang 21 Tiếng22. SĐT 980-0632034369. ^ David P. Clark; Nanette Jean Pazdernik (2012). Sinh học phân tử (tái bản lần 2). Tế bào học thuật. tr. 795. ISBN 976-0123785947. ^ Margaret C. M. Smith và R. Elizabeth Sockett, eds. (1999). Phương pháp di truyền cho sinh vật nhân sơ đa dạng . Các phương pháp trong Vi sinh vật học, tập. 29. Báo chí học thuật. tr 75 757777. Sê-ri 980-0-12-652340-9. CS1 duy trì: Sử dụng tham số sửa đổi và biên tập viên (link) ^ Morgan, Kendall. "Plasmids 101: Nguồn gốc của sự việc sao chép". addgene.org . ^ a b Russell, David W.; Sambrook, Joseph (2001). Nhân bản vô tính phân tử: hướng dẫn sử dụng trong phòng thí nghiệm . Cảng suối nước lạnh, N.Y: Phòng thí nghiệm cảng suối nước lạnh. ^ Oliveira, Pedro H.; Prather, Kristala Jones; Prazeres, Duarte M. F.; Monteiro, Gabriel A. (2010-08-01). "Phân tích lặp lại DNA trong những plasmid vi khuẩn đã cho tất cả chúng ta biết tiềm năng cho những sự kiện mất ổn định tái phát". Ứng dụng vi sinh và công nghệ tiên tiến sinh học . 87 (6): 2157 Vang2167. doi: 10.1007 / s00253-010-2671-7. ISSN 1432-0614. ^ Gonçalves, Geisa A. L.; Oliveira, Pedro H.; Gomes, Ana G.; Prather, Kristala L. J.; Lewis, Leslie A.; Prazeres, Duarte M. F.; Monteiro, Gabriel A. (2014-08-01). "Bằng chứng là những sự kiện chèn của chuyển vị IS2 bị thiên lệch về sự dịch chuyển thành phần đột ngột trong DNA đích và được điều chỉnh bởi một tập hợp những tham số nuôi cấy rất khác nhau". Ứng dụng vi sinh và công nghệ tiên tiến sinh học . 98 (15): 6609 Ảo6619. doi: 10.1007 / s00253-014-5695-6. ISSN 1432-0614. ^ Mairhofer, Juergen; Oliveira, Pedro H. (2013-09-01). "Các plasmid không còn chất đánh dấu cho những ứng dụng công nghệ tiên tiến sinh học – ý nghĩa và quan điểm". Xu hướng trong công nghệ tiên tiến sinh học . 31 (9): 539 Ảo547. doi: 10.1016 / j.tibtech.2013.06.001. ISSN 0167-7799. ^ Monteiro, Gabriel A.; Prazeres, Duarte M. F.; Prather, Kristala Jones; Oliveira, Pedro H. (2009-09-01). "Sự mất ổn định về cấu trúc của dược phẩm sinh học plasmid: những thách thức và ý nghĩa". Xu hướng trong công nghệ tiên tiến sinh học . 27 (9): 503 Ảo511. doi: 10.1016 / j.tibtech.2009.06.004. ISSN 0167-7799. PMID 19656584. ^ Uldis N. Streips, Ronald E. Yasbin, sửa đổi và biên tập. (2002). Di truyền vi sinh vật tân tiến (tái bản lần 2). Wiley-Blackwell. tr. 248. ISBN 976-0471386650. CS1 duy trì: Sử dụng tham số sửa đổi và biên tập viên (link) ^ Andrew Preston (2003). "Chương 2 – Chọn một Vector nhân bản". Ở Nicola Casali, Andrew Preston. E. Coli Plasmid vectơ: Phương pháp và ứng dụng . Các phương pháp trong Sinh học phân tử, Tập. 235. Báo chí Humana. trang 19 Tiếng26. ISBN 97-1-58829-151-6. CS1 duy trì: Sử dụng tham số sửa đổi và biên tập viên (link) ^ Kandavelou K, Chandrasegaran S (2008). "Plasmid cho liệu pháp gen". Plasmid: Nghiên cứu hiện tại và xu hướng tương lai . Caister Báo chí học thuật. Sê-ri 980-1-904455-35-6. ^ Morange M (2009). "Lịch sử cho tất cả chúng ta biết XIX. Khái niệm về tình tiết" (PDF) . Tạp chí khoa học viên học . 34 (6): 845 Cách8. doi: 10.1007 / s12038-009-0098-z. PMID 20093737. (Yêu cầu đăng ký ( giúp sức )) . ]Comptes Rendus de l'Académie des Sciences de Paris247 (1): 154–156, PMID 13561654 ^ Hayes, W (1969). "What are episomes and plasmids?". In Gordon E. W. Wolstenholme; Maeve O'Connor. Bacterial Episomes and Plasmids (CIBA Foundation Symposium). pp. 4–8. ISBN 978-0700014057. ^ Gordon E. W. Wolstenholme; Maeve O'Connor, eds. (1969). Bacterial Episomes and Plasmids (CIBA Foundation Symposium). pp. 244–245. ISBN 978-0700014057. ^ T. A. Brown (2011). Introduction to Genetics: A Molecular Approach. Garland Science. tr. 238. ISBN 978-0815365099. ^ Kathleen Van Craenenbroeck, Peter Vanhoenacker and Guy Haegeman (2000). "Episomal vectors for gene expression in mammalian cells". Eur. J. Biochem. 267 (18): 5665–5678. doi:10.1046/j.1432-1327.2000.01645.x. PMID 10971576.CS1 maint: Uses authors parameter (link) ^ Colosimo A1, Goncz KK, Holmes AR, Kunzelmann K, Novelli G, Malone RW, Bennett MJ, Gruenert DC. (2000). "Transfer and expression of foreign genes in mammalian cells" (PDF). BioTechniques. 29 (2): 314–8, 320–2, 324 passim. doi:10.2144/00292rv01. PMID 10948433. Archived from the original (PDF) on 24 July 2011.CS1 maint: Multiple names: authors list (link) ^ Gerdes K, Rasmussen PB, Molin S (1986). "Unique type of plasmid maintenance function: postsegregational killing of plasmid-không lấy phí cells". Proc. Natl. Học viện Khoa học U.S.A. 83 (10): 3116–20. Bibcode:1986PNAS…83.3116G. doi:10.1073/pnas.83.10.3116. PMC 323463. PMID 3517851. ^ Kroll J, Klinter S, Schneider C, Voß I, Steinbüchel A (2010). "Plasmid addiction systems: perspectives and applications in biotechnology". Microb. Biotechnol. 3 (6): 634–657. doi:10.1111/j.1751-7915.2010.00170.x. PMC 3815339. PMID 21255361. ^ Gunge, N; Murata, K; Sakaguchi, K (July 1982). "Transformation of Saccharomyces cerevisiae with linear DNA killer plasmids from Kluyveromyces lactis". Journal of Bacteriology. 151 (1): 462–4. PMC 220260. PMID 7045080. ^ "Vector NTI feedback video". The DNA Lab.
Further reading[edit]
- Klein, Donald W.; Prescott, Lansing M.; Harley, John (1999). Microbiology. Boston: WCB/McGraw-Hill.
Smith, Christopher U. M. Elements of Molecular Neurobiology. Wiley. pp. 101, 111.
Albert G. Moat; John W. Foster; Michael P. Spector (2002). Microbial Physiology. Wiley-Liss. ISBN 978-0-471-39483-9.
Episomes[edit]
- Piechaczek C, Fetzer C, Baiker A, Bode J, Lipps HJ (1999). "A vector based on the SV40 origin of replication and chromosomal S/MARs replicates episomally in CHO cells". Nucleic Acids Res. 27 (2): 426–428. doi:10.1093/nar/27.2.426. PMC 148196. PMID 9862961.
Bode J; Fetzer CP; Nehlsen K; Scinteie M; Hinrichsen B-H; Baiker A; Piechazcek C; Benham C; Lipps HJ (2001). "The Hitchhiking principle: Optimizing episomal vectors for the use in gene therapy and biotechnology" (PDF). Gene Ther Mol Biol. 6: 33–46. Archived from the original (PDF) on 30 May 2009.
Nehlsen K, Broll S, Bode J (2006). "Replicating minicircles: Generation of nonviral episomes for the efficient modification of dividing cells" (PDF). Gene Ther Mol Biol. 10: 233–244. Archived from the original (PDF) on 30 May 2009.
Ehrhardt A, Haase R, Schepers A, Deutsch MJ, Lipps HJ, Baiker A (2008). "Episomal vectors for gene therapy". Curr Gene Ther. 8 (3): 147–161. doi:10.2174/156652308784746440. PMID 18537590. Archived from the original on 26 September 2011.
Argyros O, Wong SP, Niceta M, Waddington SN, Howe SJ, Coutelle C, Miller AD, Harbottle RP (2008). "Persistent episomal transgene expression in liver following delivery of a scaffold/matrix attachment region containing non-viral vector". Gene Therapy. 15 (24): 1593–1605. doi:10.1038/gt.2008.113. PMID 18633447.
Wong SP, Argyros O, Coutelle C, Harbottle RP (2009). "Strategies for the episomal modification of cells". Current Opinion in Molecular Therapeutics. 11 (4): 433–441. PMID 19649988. Archived from the original on 17 September 2011.
Haase R, Argyros O, Wong SP, Harbottle RP, Lipps HJ, Ogris M, Magnusson T, Vizoso Pinto MG, Haas J, Baiker A (2010). "pEPito: a significantly improved non-viral episomal expression vector for mammalian cells" (PDF). BMC Biotechnol. 10: 433–441. doi:10.1186/1472-6750-10-20. PMC 2847955. PMID 20230618.
External links[edit]

Post a Comment